本文主要是介绍ISME | 叶片细菌群落中维生素营养缺陷的代谢适应,希望对大家解决编程问题提供一定的参考价值,需要的开发者们随着小编来一起学习吧!
点击蓝字 关注我们
Mid-Autumn Festival

编辑:朱松梅 校对:祝新宇
论文ID
原名:Metabolic adaptation to vitamin auxotrophy by leaf-associated bacteria
译名: 叶片细菌群落中维生素营养缺陷的代谢适应
第一作者:Birgitta Ryback
作者单位:Institute of Microbiology, ETH Zurich
期刊:The ISME Journal
发表时间:2022


摘要✦
营养缺陷型微生物无法合成其新陈代谢所必需的所有代谢物,需要依靠外源提供。已有大量研究对在实验室产生和进化突变的营养缺陷型菌株进行了探索,但尚未有研究系统地阐述营养缺陷型产生的适应机制。作者从拟南芥叶子中发现多达一半菌株对生物素、烟酸、泛酸和/或硫胺素有生长需求。随后作者探讨了微生物的基因型和营养缺陷型的相关性。研究结果发现营养缺陷型菌株可以储存辅酶,并通常能够在不补充维生素的情况下增长1-3倍。其中,更有菌株可以在胞体内储存生长9倍所需的生物素。在共培养实验中,并发现营养缺陷型菌株在外源添加维生素后比原始营养缺陷型菌株有更高的物种丰富度。消费者-资源模型的扩展预测了营养缺陷型菌株可以利用其他生物提供的碳水化合物,表明营养缺陷型菌株能从维生素以外的代谢副产物中获益。
背景介绍✦
辅酶对细胞代谢至关重要,它们构成许多酶催化反应的核心(如:氧化还原反应、氨基转移和C-C键形成),并且作为一碳单元和有机酸的载体。生物合成缺陷的生物体-营养缺陷型生物,必须通过外部供给获得辅酶或辅酶前体分子,如外源维生素。因为它们依赖环境中供应的维生素来促进生长,辅酶的不稳定性会加剧外部供应的需求。近期研究表明,辅酶的碳链在体内是稳定的。事实上,辅酶寿命是一种通过进化选择出的内在特性,可以认为是微生物形成营养缺陷增殖的先决条件。然而,目前尚不清楚大多数环境中细菌营养缺陷型的常见程度。
营养缺陷型微生物需要外部供应维生素,这引发了营养缺陷型菌株的生理或其他适应性问题,包括维生素存储。众所周知,营养缺陷型乳酸菌在去除维生素的情况下仍能保持生长。尽管已经证明了元素化合物(如:C、P、N)的细胞内存储,但小分子(包括维他命和辅酶)的存储目前还没有得到很好的证实。环境细菌中的储存范围有多广也尚不清楚。在参与辅酶生物合成的基因中发生突变的原养大肠杆菌菌株,即“人工”营养缺陷型突变体,在去除必需维生素后仅具有约一倍增殖潜力。这种双重过量的辅酶在细胞分裂过程中保持旺盛的新陈代谢以适应细胞的扩张的最低要求。基于这些结果,作者假设,对于自然界中发现的辅酶供应可能不稳定的营养缺陷型微生物,可能进化出了不同的策略来应对营养依赖和辅酶储存。一种化合物生物合成的缺失也可能以更系统的方式影响细胞功能。在大肠杆菌、枯草芽孢杆菌和见氏不动杆菌等模式生物中研究了生物合成丧失导致的直接生理后果,包括适合度效益和交互共生潜力。这些研究表明,只要能获得所需的营养物质,营养缺陷型菌株就能战胜野生型菌株。可以预见的是,如果有足够的进化时间,可能会选择其他特征来弥补生物合成能力的缺失。
本文首先考察了At-LSPHERE集合中营养缺陷型的现状,该集合由从自然生长的拟南芥叶子中分离出的224个菌株组成。这些菌株在富含维生素和氨基酸的培养基中分离出来,可以通过系统的营养缺失实验来检查菌株集合中的营养缺陷。然后,讨论了营养缺陷型菌株是否比原营养型菌株能保持更高的辅酶储存,并探索与营养缺陷型共同发生(并可能共同进化)的基因组特征。最后,在共培养实验中检验了维生素的获取以及营养缺陷型菌株的适应性。
结果与讨论✦
01
营养缺陷型菌株中的维生素储存
为确定本研究的营养缺陷型菌株,作者筛选了一个具有代表性的224个菌株集合(At-LSPHERE)。简而言之,在96孔板中培养菌株并做3个重复,培养基分别做如下处理:添加维生素、氨基酸、两种营养物质都添加以及不添加营养物质。在培养基中生长的156株菌株,有50%(78株)的生长需要补充,因此可能是营养缺陷型。从这78个菌株中,选择50个做进一步鉴定,发现大多数维生素营养缺陷型是针对生物素、硫胺素、烟酸和泛酸的,鉴定了针对每种维生素的超过10个营养缺陷型菌株(图1. a)。
为了克服读板器的线性读取范围的限制,研究进一步分析了22株在液体培养基中生长时不会聚团的菌株在0-(1×109/ml)之间生长速率。最终,证实这22种菌株均为营养缺陷型(图1. c)。具体来说,每个菌株平均对两种维生素有营养缺陷,80%的菌株对多种维生素有营养缺陷。
为了评估所选营养缺陷型菌株中细胞内维生素储存的潜力,用半连续生长数据计算了每株菌株在维生素缺乏后偏离指数生长之前的倍增总数,并通过与补充的对照培养物进行比较来确定。(图1. b)。因此,这些值可以作为维生素储存的指标。研究发现大多数菌株的维生素储存量在1-2倍,类似于大肠杆菌突变体。还观察到一些营养缺陷型菌株在去除生物素后的很长一段时间内保持指数增长,可达到4-5倍。金黄杆菌 Leaf201菌株可以在维生素撤出后仍然增加9倍(图1. d)。
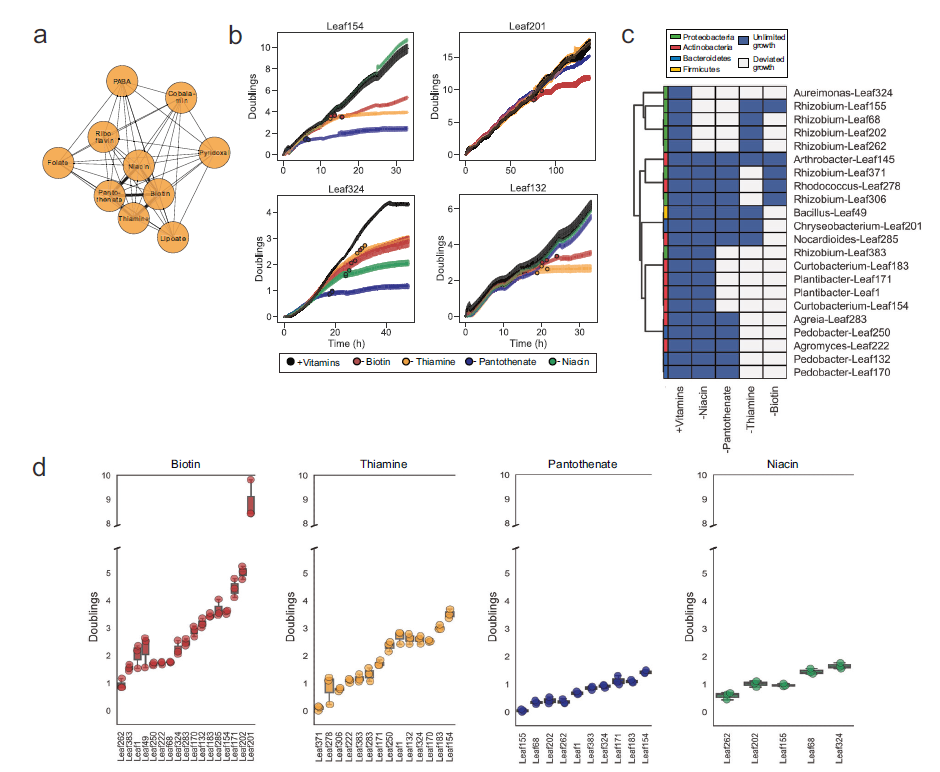
图1. 营养缺陷型菌株中维生素的储存
02
基因缺失导致营养缺陷
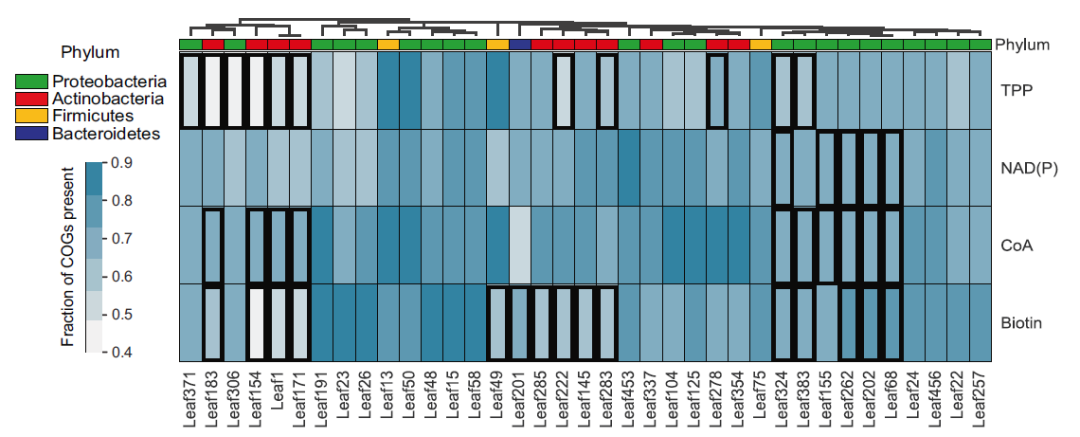
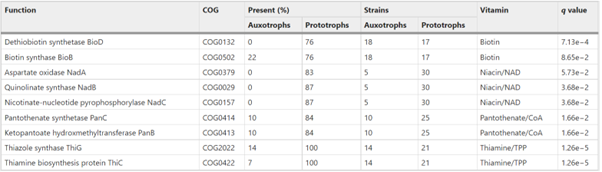
在了解了22个验证菌株对维生素的需求后,作者以13种原营养型菌株作为对照(图2. A),研究了营养缺陷型型菌株基因组中的维生素生物合成途径。所有菌株,包括原营养型菌株,在所研究的四种辅酶(生物素、硫胺素、辅酶A、NAD)的合成途径中缺少许多生物合成步骤,在营养缺陷型中也没有显著地观察到更少的步骤(图2 B)。通过对单个维生素合成基因在营养缺陷型和原营养型菌株的卡方检验,文章分别选取了1-3个基因作为营养缺陷微生物的标志性基因缺失(表1)。
A. 原营养型菌株的确定(原Supplementary Fig. 4)
B. 识别营养缺陷背后的突变
图2. 基因缺失导致营养缺陷
表1. 维生素营养缺陷微生物缺失的标志性基因
03
营养缺陷型菌株的基因组减小主要是非特异性的
本研究结果指出,营养缺陷型菌株的基因组较原营养型微生物普遍减小(图3),但是减小的幅度远超过对应的生物合成基因的缺失。
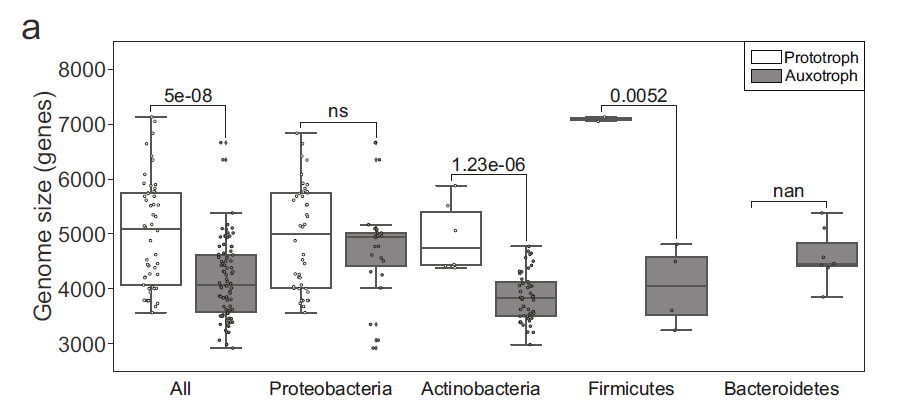
图3. 营养缺陷型与原营养型微生物基因组大小
通过对基因组建立代谢模型,作者发现对于营养缺陷型菌株与原营养型菌株不存在显著差异。因此在营养缺陷型菌株中观察到的基因组减少不能系统地解释为需要相应辅酶合成路径的减少。
为了寻找与营养缺陷相关的基因,作者分析了基因组中全部功能基在营养缺陷型和原营养型之间分布的差异(图4)。总体来说,营养缺陷型和原营养型之间的功能差异主要体现在营养缺陷型菌株基因组中功能基因的缺失。结果发现,营养缺陷型菌株在能量代谢、氨基酸合成和其他辅酶合成方面与原营养型菌株存在显著差异。
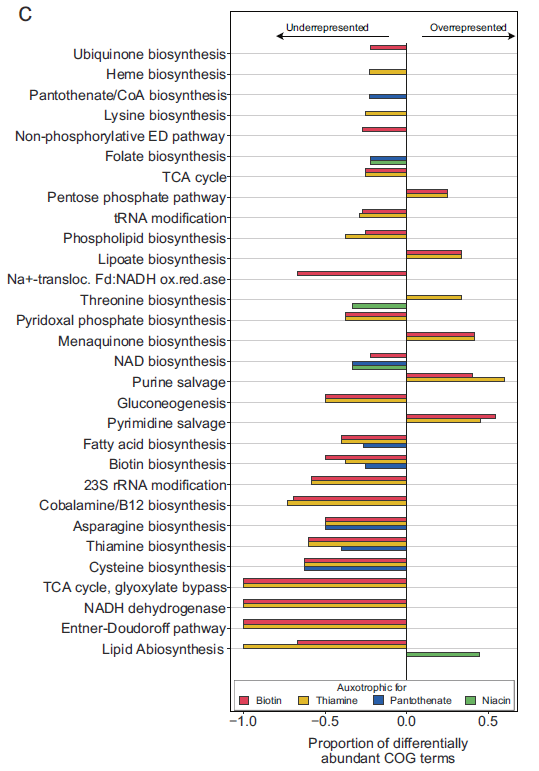
图4. 原养性和营养缺陷型菌株的全基因组比较分析
04
营养缺陷型的维生素补充来自共培养物种
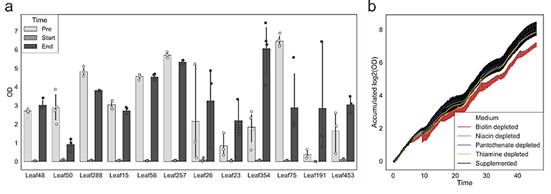
为研究营养缺陷型菌株是否从共培养的原营养型中获取维生素,作者选择了具有系统发育代表性的维生素营养缺陷型(n = 10)和原营养型(n = 10)菌株进行共培养实验(图5. a b)。通过16S rRNA基因测序对培养物进行定期分析,以确定细菌种类的相对组成。两种条件下的细菌增长是相当的(图5. c)。观察到原营养型和营养缺陷型培养基之间的相对丰度分布是相似的,并且与是否补充维生素无关(图5. d)。在具有原养菌株的群落中培养时,缺乏外部维生素补充不会阻止营养缺陷型菌株的生长,并且可能存在维生素交叉喂养。
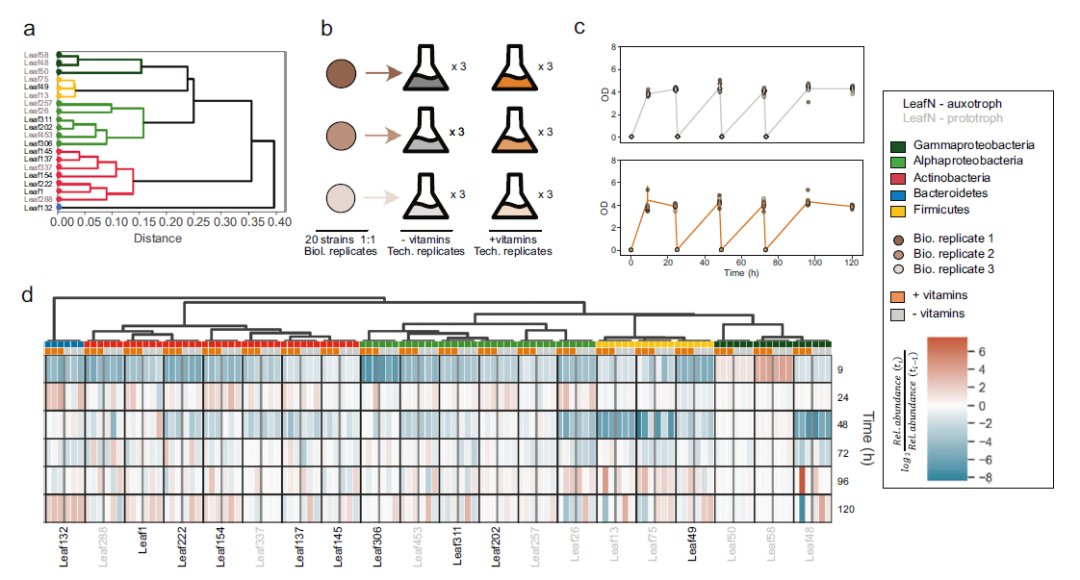
图5. 共培养实验,以确定营养缺陷型菌株是否能够从原营养体中获取维生素
通过群落胞外维生素的测定,作者发现在提供维生素的培养基中烟酸、泛酸和硫胺素0-9和9-24小时显著下降,证实了这些维生素被微生物群落吸收。但是生物素的含量没有下降,可能由于原营养型微生物分泌生物素的速度可能大于生物素营养缺陷微生物的吸收速率。在作者在培养的过程中还观测到了显著的吡哆醛的吸收,维生素B2的积累。在没有提供维生素的培养基中,作者在9h观测到了显著的烟酸、泛酸和吡哆醛的积累并在9-24 h观测到了含量的下降(图6)。
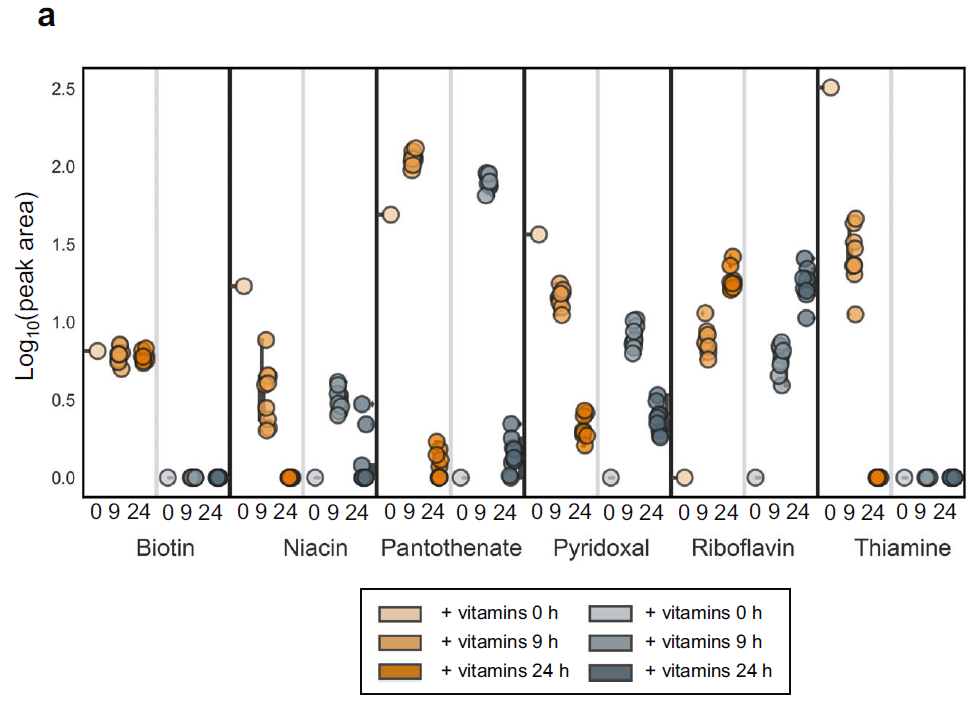
图6. 在有无维生素供给的环境中胞外维生素浓度
05
除了维生素供应,营养缺陷型菌株还受益于共培养
通过比较营养缺陷型和原营养型两组的物种丰富度随时间的变化,对添加维生素的最小培养基的相对丰度数据进行分析(图4. d),发现原营养型组的菌株明显少于营养缺陷组(图7. a)。作者通过生长速率(图7. b)和底物消耗速率与消费者-资源模型耦合,并通过共培养实验数据进行模型训练。结果发现,拓展模型仅能有效预测原营养型菌株的丰富度,并无法有效预测营养缺陷型物种丰富度的差异(图7. c)。在此基础上,作者还通过引入维生素交互喂养,碳源中间产物对模型进行拓展,以及营养缺陷微生物对于碳源中间产物的亲和力,进行了模型拓展。发现在引入以上参数后,模型可以对实验数据有较好的预测效果(图7 f)。综上所述,该分析表明,碳源交叉喂养能力的提高决定了营养缺陷型菌株在共培养中的成功。
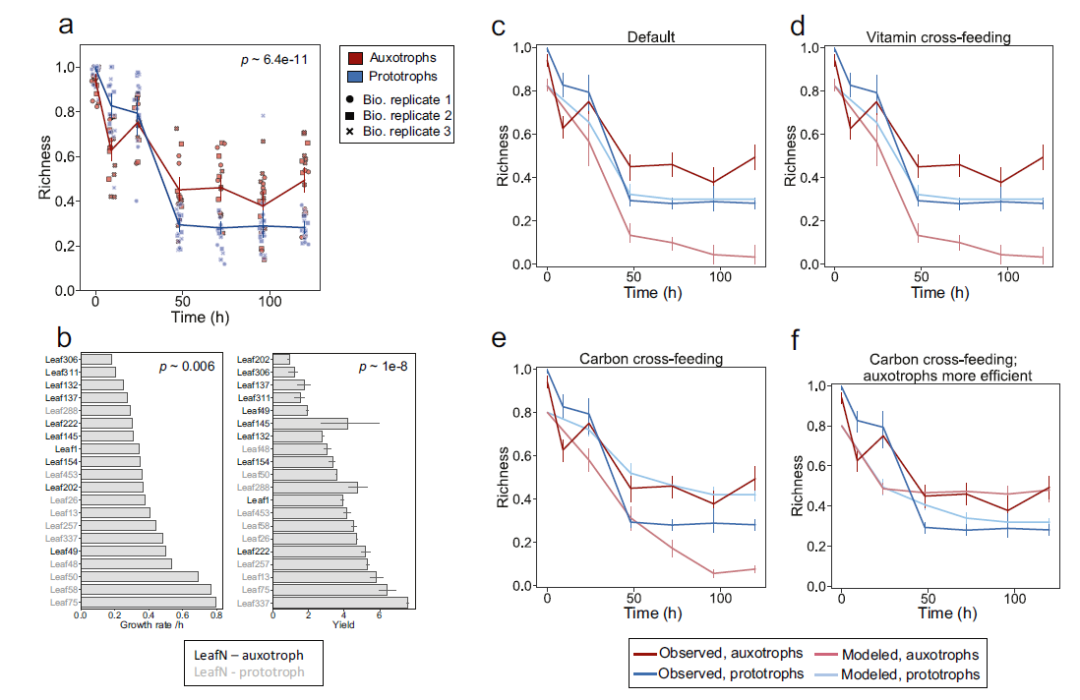
图7. 除了维生素供应,营养缺陷型菌株还受益于共培养
总结✦
营养缺陷型在进化过程中经常出现,在本研究中,作者在拟南芥叶片微生物群落内的所有主要门中发现了营养缺陷型。此外,与其他研究一致的是营养缺陷型即使在与原营养型的无维生素共培养物中也持续存在。研究证实,缺陷型菌株在补充维生素的共培养中比在单个培养中的生长更好。通过实验获得的生长速率和产量数据参数化的消费者-资源模型以及碳交叉喂养充分捕捉到了这一观察结果。由于营养缺陷型菌株在原养菌分泌足够的维生素之前无法恢复生长,因此优先使用与维生素一起分泌的碳源,可以使营养缺陷型菌株具有竞争优势。通过在消费者-资源模型框架中实施非特定的碳交叉喂养来解释稳定共存,并且这种代谢副产物的生长通常与葡萄糖等主要碳源的生长相当。消费者-资源模型框架表明,这种代谢促进作用不仅促进了共存,而且对碳副产物的优先消费也可能是一种避免竞争排斥的可行策略。总之,营养缺陷型是一种生活方式的一部分,专门消耗其他细菌的代谢产物,因此有利于作为部分微生物群落的自由生活细菌。

点击下方“阅读原文”查看更多
*:本文如有不足,请批评指正
1
END
1
EMBLab
环境微生物组与生物技术实验室
环境微生物组学研究环境中全部微生物及其遗传信息,其方法学基础与理论拓展应用是国际学术前沿和热点。西湖大学环境微生物组与生物技术实验室开展环境工程学与微生物学交叉学科研究,研究兴趣包括:1)环境工程与合成微生物组学;2)微生物组的群落构建理论与功能原理;3)抗生素耐药性产生与传播机制及风险监控;4)持久性有机污染物(塑料与药物)降解转化及健康效应。
微信号|envmbio
实验室网站|http://www.ju-emblab.com

EMBLab成员
实验室目前招聘:
博士后 2 名:塑料生物降解与酶资源发掘方向、环境微生物组与耐药组研究方向各1名。
科研助理2名:污水处理与资源化、微生物组或生物信息学方向各1名。
欢迎大家与我们联系。
联系邮箱:
jufeng@westlake.edu.cn
xuyisong@westlake.edu.cn

鞠峰
EMBLab负责人
鞠峰,西湖大学研究员、博士生导师,环境微生物组与生物技术实验室负责人,浙江省海岸带环境与资源研究重点实验室副主任。2015年获香港大学工学博士学位,之后在瑞士联邦水科学与技术研究所 (EAWAG) 从事博士后研究,2018年至今在西湖大学担任特聘研究员,开展环境微生物组科学与工程技术研究。目前担任中国工程院院刊Engineering、Frontiers in Microbiology、Engineering in Life Science 等SCI期刊编委,以及 Critical Reviews in Environmental Science Technology、Journal of Environmental Sciences、Environmental Science & Ecotechnology、The Innovation、iMeta 等期刊青年编委。曾担任加拿大自然科学与工程理事会(NSERC)国际评审专家。曾获中国工程院院刊 Engineering“编委年度贡献奖”(2021)、中国生态学会“水云天微生物生态青年科技创新奖-特等奖”(2018)、香港科学会“青年科学家奖”(2016)、香港大学“杰出研究生奖”(2015)等科研奖项。近三年主持或参与国家级或省部级科研项目 4 项;目前参编中英文专著 4 本,在 Nature Communications、ISME Journal、Advanced Science、Microbiome 、Environmental Science & Technology、Water Research 等知名学术期刊发表论文60余篇,引用 4000次。
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
系列教程:微生物组入门 Biostar 微生物组 宏基因组
专业技能:学术图表 高分文章 生信宝典 不可或缺的人
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
扩增子分析:图表解读 分析流程 统计绘图
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文
这篇关于ISME | 叶片细菌群落中维生素营养缺陷的代谢适应的文章就介绍到这儿,希望我们推荐的文章对编程师们有所帮助!







