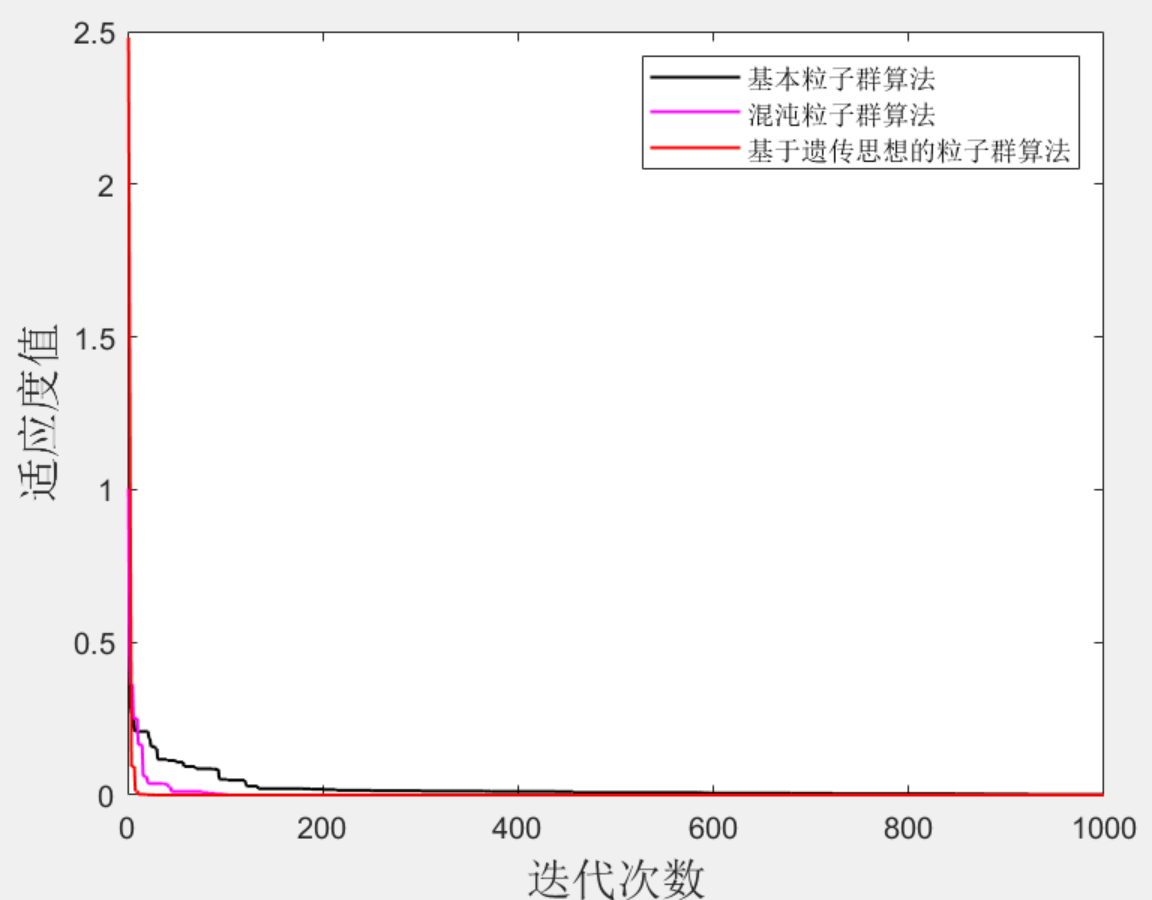
来自知乎:https://zhuanlan.zhihu.com/p/51355201
2018 年 7 月 6 日,表观遗传领域的开拓者之一 Danny Reinberg 在 Science 发表了观点(Perspectives)文章[4],就“哪些组蛋白修饰可以遗传及其机制”进行了探讨。以下对该文的核心观点进行了梳理。
染色质的两种状态
未修饰的组蛋白带正电,而 DNA 带负电,两者通过静电作用结合在一起形成核小体;组蛋白与邻近的核小体之间也具有静电作用,因此在默认情况下,染色质应该是致密压缩的(异染色 质,heterochromatin),基因整体上处于抑制状态。而无论是DNA 的复制、转录还是修复,都需要染色质处于开放状态(常染色质,euchromatin),那么问题来了,染色质是怎么打开的呢?
本内容来自:Nat Rev Genet | 20 年来表观遗传领域核心进展之全景解读 - xupenggoing的文章 - 知乎 https://zhuanlan.zhihu.com/p/57700931
表观遗传学的基础:
染色质状态包括基因处于活性状态的常染色质(euchromatin)和基因处于抑制状态的异染色质(heterochromatin)
在同一细胞核中,相同的遗传物质可以保持不同的“开”或“关”的状态,但其背后的机制所知甚少。
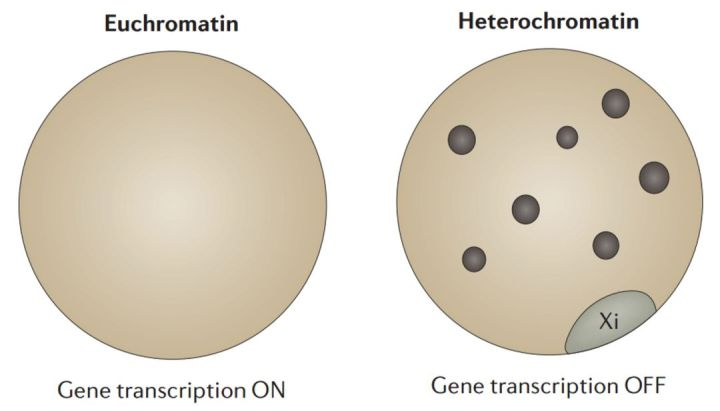
图 1. 细胞学上可见的活性染色质(常染色质)和抑制性染色质(异染色质)状态。图中显示了两个来源于雄鼠体细胞的处于细胞间期的细胞核,左边的细胞核中的 DNA 呈现出广阔的非致密的染色,而右边的细胞核呈现出具有典型的异染色质核(黑色的点),而且通过 DAPI 染色,AT 富集的重复序列可见。此外,在核周围的致密染色的 Barr 体(一个失活的 X 染色体)被单独标注出来。
2.1 DNA 甲基化
早在 1948 年,DNA 碱基的化学修饰就已被检测到(编者注:这早于1953年DNA双螺旋结构的发现);在 20 世纪 70 年代中期,Holiday 和 Pugh 就提出了 DNA 甲基化(特别是 5-甲基胞嘧啶)在基因调控中的作用。到了 1980 年,DNA 甲基化和基因抑制的联系被建立,同一时期也发现了 CpG 岛。5-氮杂胞苷(也称为 2'-脱氧-5-氮杂胞苷,后称为地西他滨)是第一种“表观遗传药物”,它能够阻断 DNA 甲基化,用于改变成纤维细胞系的基因表达和表型。此后不久, Feinberg 和 Vogelstein 报道了癌症中全基因组 DNA 低甲基化;十年后,肿瘤抑制基因的局部 DNA 高甲基化也被报道出来。这些发现为 DNA 甲基化的“酶学”理论(enzymology of DNA methylation)提供了令人信服的证据。小鼠 DNA 甲基化转移酶1(DNMT1)的成功纯化与克隆,以及 Dnmt1 突变小鼠的构建和分析向着这一理论迈出了重大一步。同一时期,第一个 DNA 甲基结合蛋白 MeCP2 被鉴定出来。时至今日,DNA 甲基化和 5-甲基胞嘧啶(被认为是“第五个碱基”)已经被确立为许多生物体内关键的表观遗传学机制。