本文主要是介绍项目文章|真菌ChIP-seq+RNA-seq解析脱落酸生物合成的调控机制,希望对大家解决编程问题提供一定的参考价值,需要的开发者们随着小编来一起学习吧!
组蛋白翻译后修饰是表观遗传调控的主要机制之一,已被证明在基因表达的调控中发挥重要作用,参与真菌发育、感染相关的形态发生、环境应激反应、次级代谢产物的生物合成和致病性。我们分享过不少真菌组蛋白修饰的文章,今天接着带来一篇利用ChIP-seq解析真菌脱落酸生物合成机制的文章。

2024年3月4日,中国科学院成都生物研究所谭红研究员课题组在期刊Frontiers in Microbiology发表文章《Normal distribution of H3K9me3 occupancy co-mediated by histone methyltransferase BcDIM5 and histone deacetylase BcHda1 maintains stable ABA synthesis in Botrytis cinerea TB-31》。该研究以灰葡萄孢(Botrytis cinerea)为研究对象,发现H3K9甲基转移酶BcDIM5与H3K14去乙酰化酶BcHda1物理相关,缺失这两者会严重影响ABA合成。RNA-seq和ChIP-seq的联合分析揭示了BcDIM5和BcHda1缺失导致H3K9me3修饰的正态分布和水平出现显著的全局缺陷。研究结论认为ABA基因簇由BcDIM5和BcHda1共同调控,对B. cinerea TB-31 ABA基因簇H3K9me3的正态分布至关重要。这项工作扩展了我们对ABA生物合成复杂调控网络的理解,为高产ABA菌株的遗传改良提供了理论基础。爱基百客为该研究提供ChIP-seq的技术支持。

-
研究背景
脱落酸(ABA)是一种保守且重要的“倍半萜烯信号分子”,广泛分布在不同生物体中,具有独特的生物学功能。ABA可以协调微生物和宿主之间的互惠和竞争关系。此外,ABA还调节植物和动物的免疫和应激反应。因此,ABA在农业、医药及相关领域有着广泛的应用。作为ABA生产模型菌株,植物病原性子囊菌Botrytis cinerea已被广泛研究。然而,关于Botrytis cinerea中ABA生物合成调控机制的研究相对较少。
-
研究路线
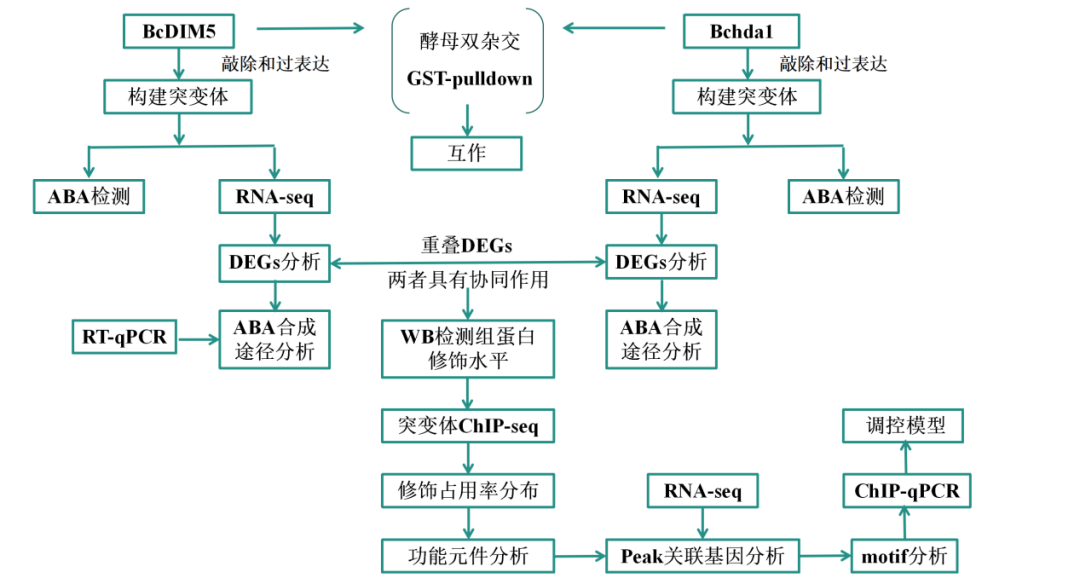
-
研究结果
-
BcDIM5是灰葡萄孢TB-31生产ABA所必需的
在之前的研究中,作者发现全局调控因子BcLae1的敲低显著降低了ABA的产生,并改变了H3K9me3的整体修饰。于是,作者推测H3K9me3修饰的变化是否对ABA的合成有影响。基于此,作者敲除了灰葡萄孢TB-31的H3K9甲基转移酶BcDIM5。研究结果表明,敲除突变体的ABA产量大幅下降(减少 97%)(图1);而在Bcdim5互补后,ABA的产量得以恢复。这些结果证明了BcDIM5参与了ABA合成的调控。
当Bcdim5被敲除时,ABA的合成从早期到后期严重减少。为了了解Bcdim5基因敲除对ABA合成相关途径的影响,作者选择了ABA合成的早期阶段进行RNA-seq分析。对差异表达基因进行分析,共获得1002个DEGs,其中上调基因769个,下调基因233个(图 1B)。
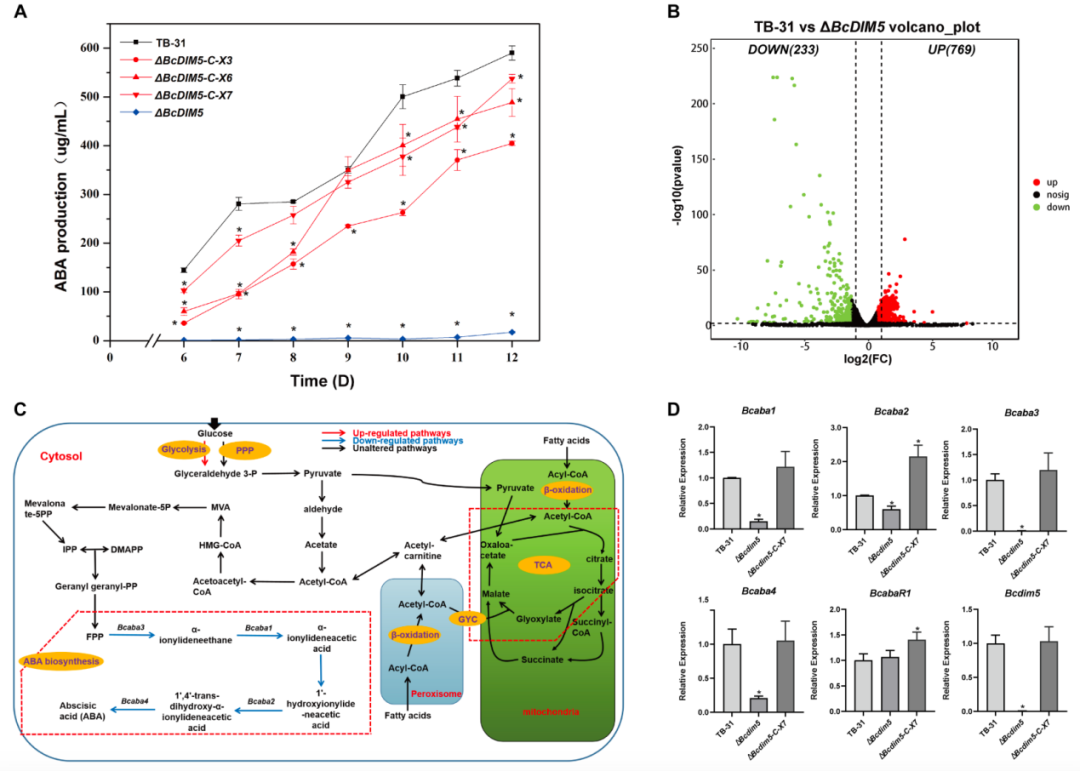
图1:灰葡萄孢TB-31生产ABA需要Bcdim5。
随后,作者专注于分析与ABA合成相关途径的表达变化,包括碳水化合物运输和利用、乙酰辅酶A合成和运输相关途径、FPP合成和ABA合成基因簇。相关基因的表达水平有减少的,也有没有变化的。基于此,作者发现ΔBcdim5中ABA产量的减少不能归因于乙酰辅酶a的合成和转运。此外,参与MVA通路的酶的转录水平保持不变,而ABA合成基因簇(Bcaba1、Bcaba2、Bcaba3和Bcaba4)的转录水平显著降低。因此,作者认为这是导致ΔBcdim5中ABA产量降低的直接原因,这一观点经RT-qPCR证实。
2. 组蛋白去乙酰化酶BcHda1与BcDIM5相互作用,参与ABA合成的调控
利用生信分析,作者推测BcHda1是BcDIM5的潜在相互作用蛋白。利用酵母双杂交、GST-pulldown证明了它们之间的互作关系。随后,作者将Bchda1敲除。突变体ΔBchda1的ABA产量与ΔBcdim5相似。同样地,利用RNA-seq分析来评估Bchda1敲低对ABA合成途径的影响。结果显示,除了ABA合成基因簇的转录水平显著下降(图2E)外,其他相关通路的表达变化都没有,这表明BcDIM5和BcHda1通过调节ABA基因簇的转录水平来影响ABA的合成。比较分析ΔBchda1和ΔBcdim5突变体的RNA-seq数据,DEGs数据的重叠表明,这两种蛋白具有协同作用,发挥调节功能。
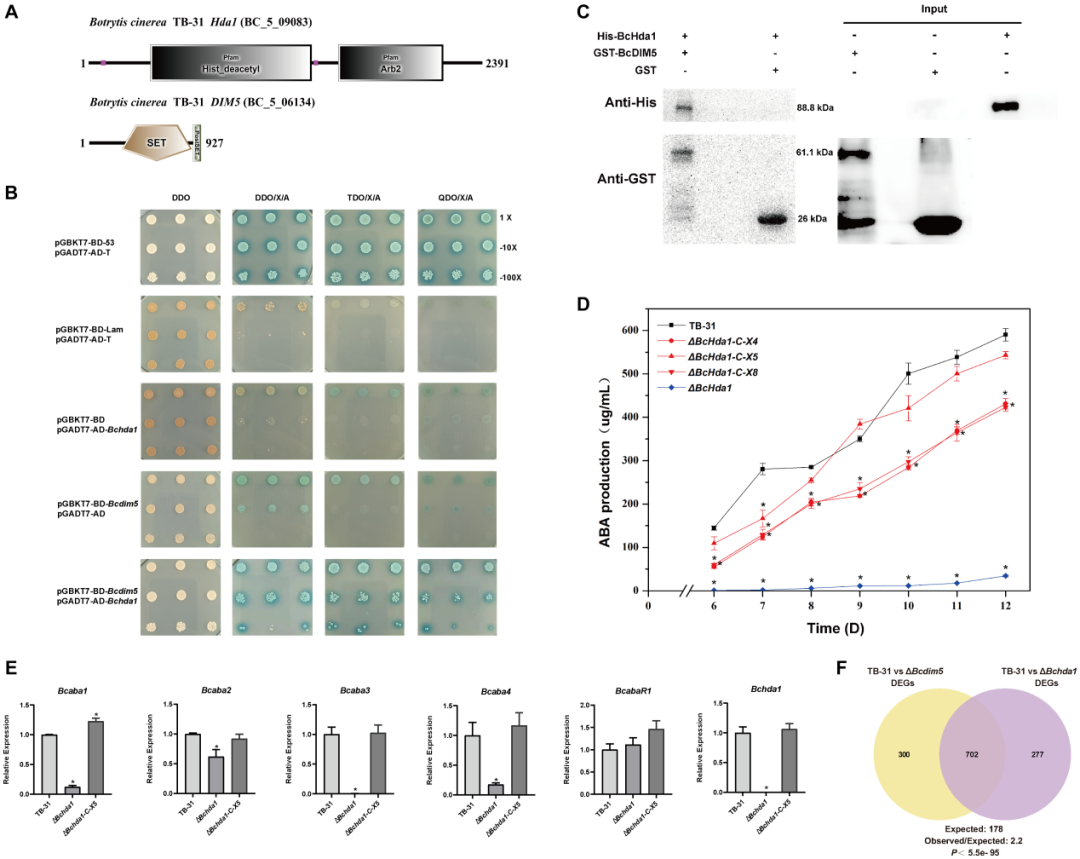
图 2:BcDIM5的相互作用蛋白BcHda1也参与了ABA的合成
3. 过表达Bcdim5和Bchda1并没有增加灰葡萄孢TB-31中ABA的产生
构建Bcdim5和Bchda1超表达突变体,并检测其ABA产量。同样,对超表达突变体进行RNA-seq分析。过表达Bcdim5和Bchda1并没有增加灰孢杆菌TB-31中ABA的产量。基于这部分的结果,作者认为Bcdim5和Bchda1的适当表达水平是维持ABA合成稳定的重要因素。
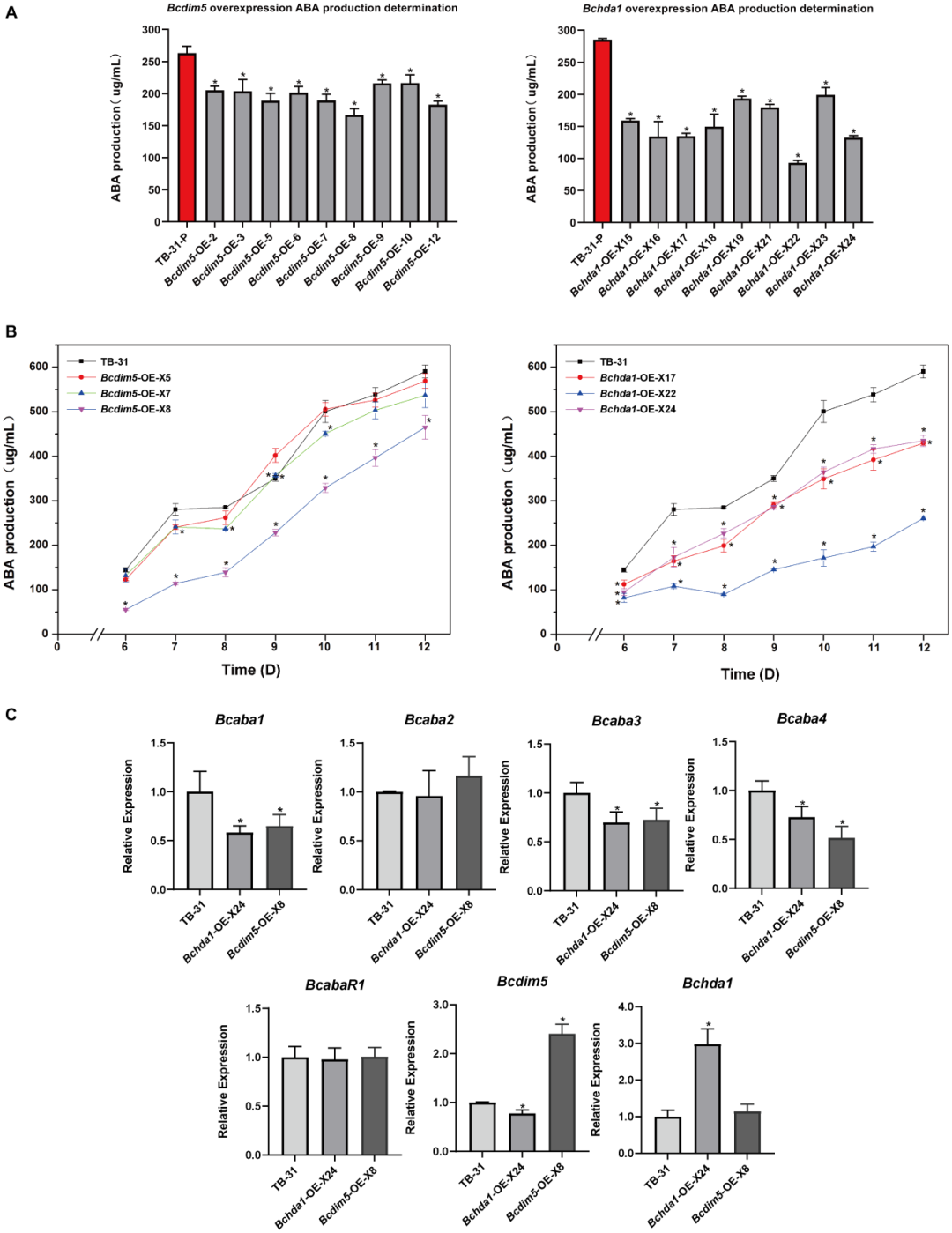
图 3:过表达Bcdim5和Bchda1并没有增加葡萄灰霉病菌TB-31中ABA的产量。
4. BcHda1和BcDIM5对于H3K9me3占用率的正态分布至关重要
由于BcDIM5是组蛋白H3K9甲基转移酶,而BcHda1可能是组蛋白H3K14去乙酰化酶,它们的缺失和过表达将不可避免地影响染色质整体修饰状态的改变。结果表明,在Bcdim5和Bchda1突变体中,H3K9me和H3K9me2没有明显变化。然而,在ΔBchda1和ΔBcdim5中,H3K9me3的修饰水平显著降低。基因互补后修饰水平的恢复表明,BcDIM5和BcHda1在调控H3K9me3的整体修饰水平中发挥了一定作用。因此,作者推测,H3K9me3整体修饰状态的改变是调控Bcdim5和Bchda1突变体共享的DEGs的主要因素。同时,研究发现Bchda1过表达导致了H3K14ac整体修饰水平的降低。结果表明,BcHda1作为组蛋白H3K14去乙酰化酶,从而影响H3K14ac的整体修饰水平(图4A,B)
H3K9me3和H3K14ac在真菌发育、毒力和次级代谢产物中发挥着至关重要的作用,前者通常与基因抑制相关,后者与基因激活相关。为了更深入地了解Bchda1和Bcdim5缺失对菌株H3K9me3和H3K14ac占用率的影响,作者通过ChIP-seq绘制了这些修饰在ΔBcdim5和ΔBchda1突变体中的基因组分布,以灰葡萄孢TB-31作为背景对照。结果显示,与TB-31相比,ΔBcdim5中的H3K14ac水平几乎没有变化。相比之下,与TB-31相比,ΔBchda1的H3K14ac占用率更高。然而,仅鉴定出105个基因对应的128个H3K14个差异峰,表明富集区占整个基因组的极小比例。这说明Bchda1的缺失对全局H3K14ac的影响相对较小。值得注意的是,与TB-31相比,ΔBchda1和ΔBcdim5突变体的H3K9me3占用率分布均存在显著不足。
ΔBchda1中998个基因和ΔBcdim5中1191个基因的H3K9me3水平至少降低了1倍。BcHda1和BcDIM5共同介导的H3K9me3去除位点分布在注释基因的不同区域,包括外显子、启动子、内含子和基因间区域,外显子和启动子的比例占比较大(图4C)。Meta-gene分析也显示ΔBcdim5和ΔBchda1菌株的H3K9me3水平下降,特别是在TSS和TES区域之间(图4D)。另外,在ΔBchda1和ΔBcdim5株中,254个基因和80个基因分别被H3K9me3占据,而在TB-31中缺失。总的来说,BcDIM5和BcHda1的缺失导致了H3K9me3修饰正态分布和水平的严重整体缺陷,包括大部分缺失丢失和减少,以及基因组位点的局部高甲基化。

图4:Bcdim5和Bchda1对于灰葡萄孢TB-31中全基因组H3K9me3占据的正态分布至关重要
5. BcHda1和BcDIM5的缺失可重编程H3K9me3占据的基因表达
为了进一步比较H3K9me3在ΔBcdim5和ΔBchda1菌株之间的分布,作者定量分析了H3K9me3占据Peak关联的基因数量。研究共鉴定出2846个H3K9me3占据峰(ΔBcdim5 vs TB-31),对应ΔBcdim5中的1964个基因;2069个峰(ΔBchda1 vs TB-31)对应ΔBchda1中的1539个基因。其中,809个基因有重叠的H3K9me3占据峰(图4E),说明这些基因的H3K9me3富集区的修饰可能受到BcHda1和BcDIM5的共同调控。随后,作者探讨了BcHda1和BcDIM5在基因抑制过程中的生物学联系。
RNA-seq分析表明,ΔBcdim5和ΔBchda1菌株的DEGs具有相同的失调方向,且上调基因(611个上调基因)存在显著重叠。同时,上调的交集中有193个基因,在ΔBcdim5和ΔBchda1菌株中缺少或减少了H3K9me3修饰,表明BcDIM5和BcHda1对这些基因的转录沉默是必需的(图4F)。此外,上调的交叉集中有158个基因与H3K9me3删除峰区域相邻,这表明H3K9me3的扩散可能对基因的抑制是必需的。这些结果表明BcDIM5和BcHda1对B. cinerea TB-31中H3K9me3介导的转录沉默至关重要。为了更深入地了解BcDIM5和BcHda1对B. cinerea TB-31的全基因组H3K9me3修饰的特征,使用HOMER软件预测和分析了H3K9me3占据峰的motif。结果显示,丰度最高的基序是GTTGTNG,其次是GTCCTC,GCCCTT和GGCGA等(图4G)。H3K9me3在这些motif上的结合可能需要BcDIM5和BcHda1的共同参与。
由BcDIM5和BcHda1引起的组蛋白修饰变化也影响了B. cinerea的次级代谢物(SM)关键酶基因的转录水平。在ΔBcdim5中,Bcstc1、Bcstc3、Bcpks11、Bcpks17和Bcpks21基因的转录水平降低,而Bcstc4、Bcpks1、Bcpks10和Bcpks14基因的转录水平增加。结合ChIP-seq分析,观察到ΔBcdim5在Bcstc3的外显子区域中H3K9me3占据增加,而在Bcpks1和Bcpks10的外显子区域中H3K9me3占据减少(图5A-D)。此外,在Bchda1-OE-X24菌株中,五个聚酮合酶Bcpks6、Bcpks9、Bcpks11、Bcpks1和Bcpks21基因的转录水平均增加。ChIP-qPCR分析显示Bcpks6和Bcpks21基因的启动子区域中H3K14ac占据增加,这可能解释了观察到的转录上调。由BcDIM5和BcHda1引起的组蛋白修饰重编程促进了B. cinerea中新型次生代谢产物的进一步挖掘。
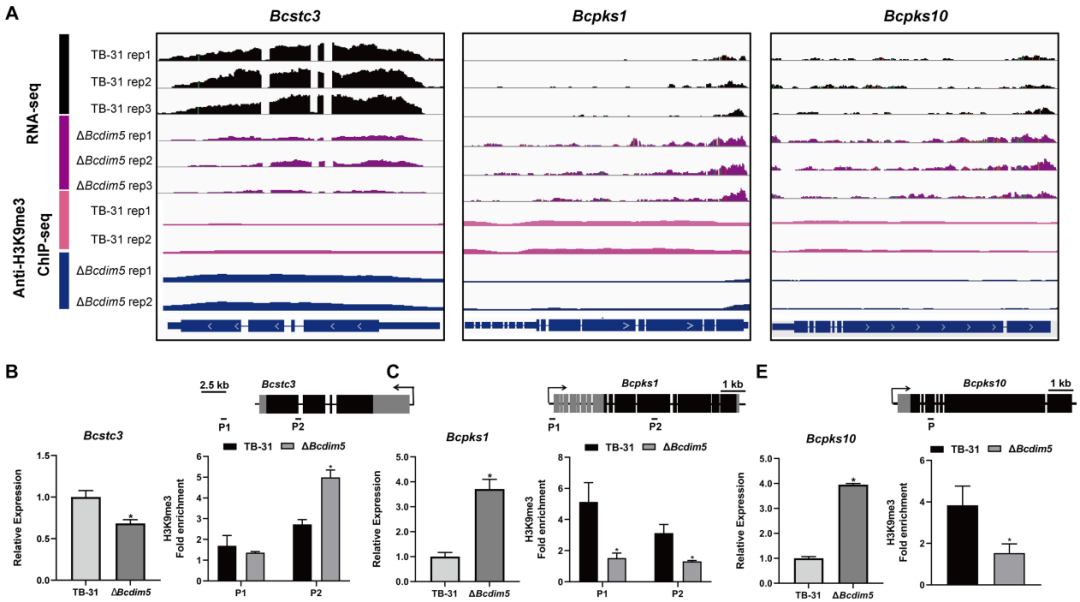
图5:由于Bcdim5确实导致差异H3K9me3占据,从而改变了SM相关基因的表达。
6. BcHda1和BcDIM5的缺失导致ABA基因簇的H3K9me3超甲基化
目前的研究表明,H3K9me3通常与生物合成基因簇的抑制有关,并被进一步确定为组成型异染色质的标记,主要与着丝粒异染色质相关。因此,在病原真菌中敲除DIM5会导致一些次生代谢产物的产量增加。然而,DIM5通常与该菌株的致病性有关。DIM5的丢失伴随着菌株生长发育缓慢、毒性减弱,毒力因子合成减少,这也发生在灰葡萄孢B. cinerea中。在ΔBcdim5和ΔBchda1突变体中,毒力因子ABA的产生严重受损。对TB-31、ΔBcdim5和ΔBchda1中ABA基因簇的H3K14ac和H3K9me3富集情况进行IGV分析,发现H3K14ac修饰无显著性差异。
出乎意料的是,Bcdim5和Bchda1的缺失并没有导致ABA基因簇中H3K9me3占用率的减少,而是导致H3K9me3的过度富集(图6A)。随后,利用ChIP-qPCR进一步验证了H3K9me3在Bcaba1和Bcaba3基因的启动子区和外显子区的占用率显著增加(图6B),这进而导致了整个基因簇的表达下调。需要进一步调查这种超级H3K9me3修饰的产生是否是由于Bcdim5和Bchda1的删除导致其他组蛋白修饰酶被招募到ABA基因簇中。然而,毫无疑问,BcDIM5和BcHda1对ABA合成是必需的。同时,作者还发现BcDIM5和BcHda1并未通过电泳迁移位移实验直接结合到ABA基因簇的启动子区域。因此,作者认为其他蛋白质协助BcDIM5和BcHda1对ABA基因簇的H3K9me3修饰(图6C)。
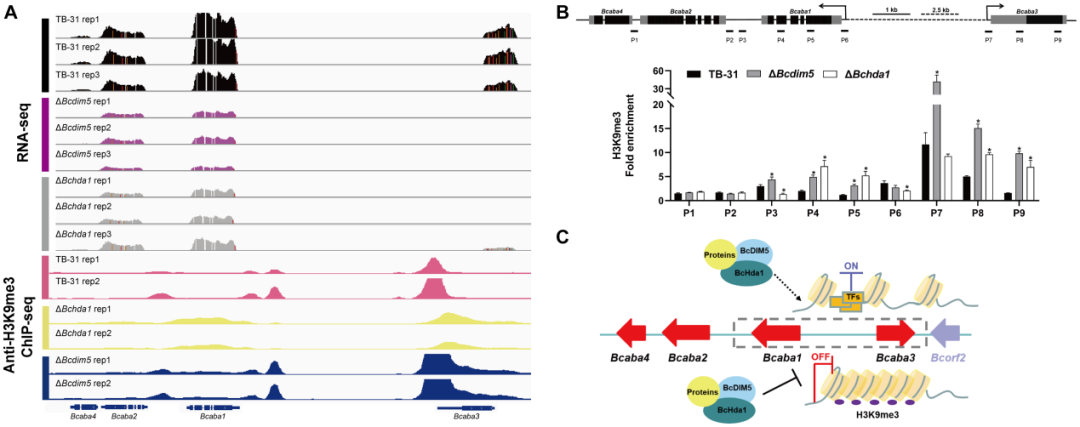
图6:ABA基因簇表达水平的降低与H3K9me3占用率的增加有关。
除了ABA基因簇外,作者还对另外两个已知的毒力因子(BOT和BOA)进行了分析。RNA-seq数据显示,ΔBchda1菌株中BOA和BOT基因簇的表达水平保持不变。然而,在ΔBcdim5菌株中,Bcboa3、Bcboa15、Bcboa16、Bcboa17以及整个BOT基因簇(Bcbot1-5)的表达水平下调。IGV显示在ΔBcdim5菌株中,Bcboa3、Bcboa15和Bcboa16基因的外显子和启动子区域中H3K9me3占据增加,这可能导致表达下调(图7A-D)。值得注意的是,BOT基因簇在H3K9me3占据上没有显示出显著变化,因此作者推测Bcdim5的敲低间接导致了整个簇的转录水平下调。Bcdim5和Bchda1的删除破坏了一些毒力因子(ABA、BOA)的正常翻译后修饰,这也反映了毒力因子调控网络的复杂性。结合以上研究分析,作者得出结论:ABA基因簇受BcDIM5和BcHda1共同调控,它们对B. cinerea TB-31 ABA基因簇H3K9me3的正态分布和维持至关重要。
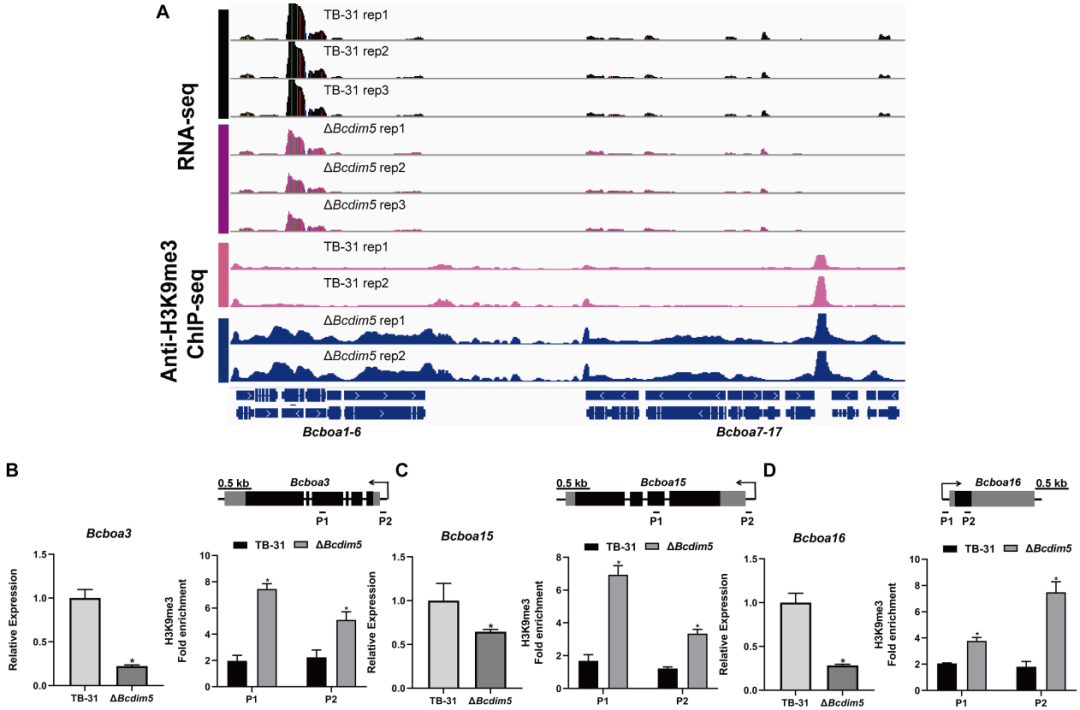
图7: BOA基因簇表达水平的降低与H3K9me3占用率的增加相关
总 结
研究结果确定了H3K9甲基转移酶BcDIM5和H3K14去乙酰化酶BcHda1之间的物理联系,揭示了它们在ABA生物合成中的调控作用。BcDIM5和BcHda1在B. cinerea TB-31中H3K9me3占据的正态分布和次生代谢产物稳定合成中发挥着关键作用。据作者所知,这是关于组蛋白修饰参与B. cinerea ABA生物合成的首次报道。这项工作为进一步改良高产ABA菌株提供了理论基础,并为在B. cinerea中发现和生产未知次生代谢产物提供了新的启示。
爱基百客拥有10年表观遗传学研究服务的经验,我们提供CUT&Tag、ATAC-seq、ChIP-seq、DAP-seq等表观遗传学测序服务。服务范围涵盖实验设计、样本处理、数据分析到验证的全流程,旨在为客户提供全面、专业的技术支持。目前,我们在微生物的表观经验已相当丰富,辅助客户多次发表Nature Commun、Plant Commun、Nucleic Acids Res、New Phytol等国际高水平期刊,欢迎有需求的老师咨询~
微生物表观经验部分汇总


这篇关于项目文章|真菌ChIP-seq+RNA-seq解析脱落酸生物合成的调控机制的文章就介绍到这儿,希望我们推荐的文章对编程师们有所帮助!






