本文主要是介绍项目文章| PBJ(IF:13.8)发表稻曲病菌效应因子Uv1809增强组蛋白去乙酰化抑制水稻免疫的分子机制,希望对大家解决编程问题提供一定的参考价值,需要的开发者们随着小编来一起学习吧!

稻曲病菌(Ustilaginoidea virens)感染引起的稻曲病(RFS)不仅严重影响水稻生产而且威胁人类健康。病原体侵染植物的过程中,植物进化出独特的先天免疫系统。已知组蛋白乙酰化相关酶参与植物免疫调控过程,那么稻曲病菌是如何通过组蛋白乙酰化影响水稻免疫的呢?

发表单位:安徽农业大学
期 刊 :Plant Biotechnology Journal(IF:13.8)
2023年安徽农业大学陈晓洋/潘月敏团队在期刊Plant Biotechnology Journal(IF:13.8)发表了题为“Ustilaginoidea virens-secreted effector Uv1809 suppresses rice immunity by enhancing OsSRT2-mediated histone deacetylation”的研究论文。该研究结果表明Uv1809是针对水稻组蛋白去乙酰基酶OsSRT2的关键效应因子,Uv1809通过促进OsSRT2介导组蛋白去乙酰化,降低宿主中H4K5ac和H4K8ac的水平,干扰防御基因转录的激活。该研究揭示了一种新的反防御机制,植物病原体通过分泌效应器来干扰宿主的表观遗传修饰,从而使宿主防御反应失活。爱基百客为该研究提供small RNA-seq和ChIP-seq的技术支持。


1 研究背景
由稻曲病菌(U.virens)感染引起的稻曲病(RFS)是影响全球水稻生产最具破坏性的病害之一。RFS不仅会降低产量,其产生的环肽霉菌毒素,威胁动物和人类健康,其中一些效应因子在感染中发挥着关键作用。另一方面,植物面对环境中植物病原体的攻击进化出了独特的先天免疫系统。植物病原体将致病效应因子分泌到宿主细胞抑制基础植物基础防御,从而感染宿主。然而,U.virens与水稻互作的分子机制在很大程度上尚不清楚。
2 技术路线
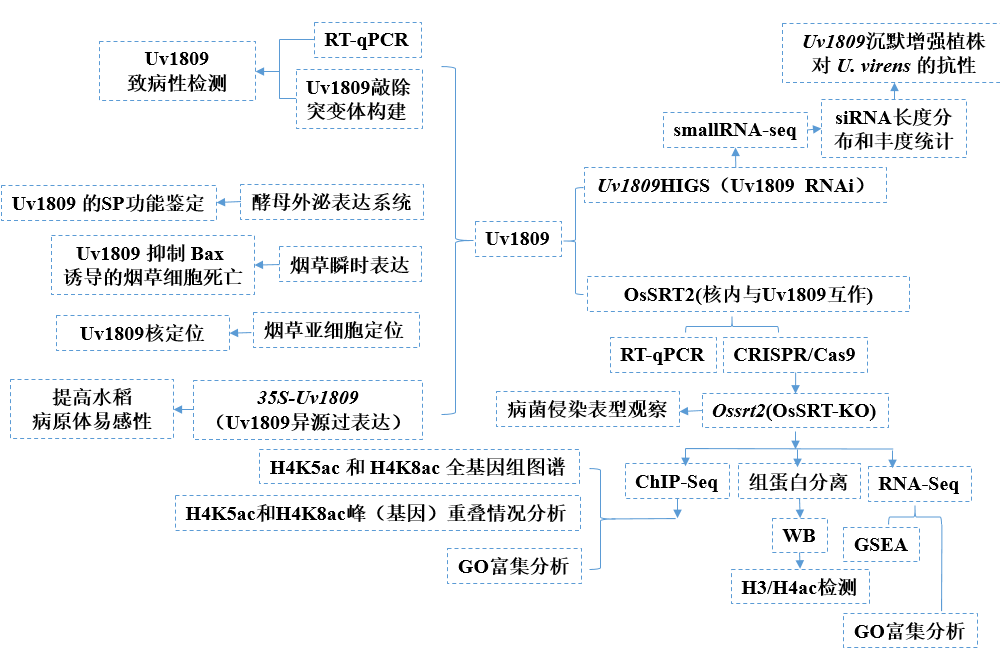
3 研究结果
1. Uv1809是U. virens完全致病所必需
U. virens感染的水稻小穗转录组数据中发现Uv1809在感染早期显著上调。RT-qPCR数据进一步证实这一结果(图1a)。Uv1809序列分析和Pfam domain预测表明Uv1809编码一个 391-aa 蛋白,包含一个信号肽(SP),并且其C 端含有大量重复序列,Uv1809功能上被注释为假定蛋白。系统发育树分析表明Uv1809同源物只在真菌中广泛分布。
之后作者利用农杆菌转化法构建Uv1809敲除突变体(ΔUv1809-1 和 ΔUv1809-3)和互补菌株CΔUv1809-1。数据显示ΔUv1809 突变体的致病性显著降低(图1b,c)。但ΔUv1809 突变体的菌丝生长速度和分生孢子量与对照组没有明显差异。Uv1809 是 U. virens 完全致病所必需的。
2. Uv1809 的信号肽对其分泌和功能至关重要
使用酵母分泌系统验证Uv1809 的SP分泌活性。编码 22-aa SP的序列与pSUC2 载体连接后转化到酵母菌株 YKT12中,检测酵母YTK12生长情况和 TTC 培养基中的转化酶活性。结果表明Uv1809SP 和Avr1bSP (阳性对照)转化株能在 YPRAA 培养基上生长,阴性对照 pSUC2 转化株和未转化 YTK12 菌株不能在 YPRAA 培养基上生长。此外,与对照相比Uv1809SP 和 Avr1bSP 转化子分泌转化酶,使 TTC 溶液变红(图 1d)。这表明Uv1809 的 SP 是一个功能性分泌信号,Uv1809 是一种分泌蛋白。
作者进一步检测Uv1809的 SP是否与U. virens致病性相关。CΔUv1809ΔSP-1 转化株(缺失SP的Uv1809(Uv1809ΔSP)导入ΔUv1809-1突变体)显示出与ΔUv1809-1菌株相似的致病性,与对照相比,致病性显著降低(图 1b,c)。因此,Uv1809 的 SP 在 U. virens 的致病性中起着关键作用。
3. Uv1809 可抑制 Bax 诱导的烟草细胞死亡
为了明确Uv1809在宿主免疫中的作用。作者构建pVX-Uv1809并导入农杆菌EHA105中。使用该农杆菌浸润烟草叶片,未观察到细胞死亡(图 1e)。相同条件下使用小鼠促凋亡蛋白Bax表达的农杆菌浸润烟草,观察到细胞死亡。但Uv1809 和 Bax 蛋白共表达,Bax 诱导的细胞死亡被抑制(图 1e)。为了明确 Uv1809 内介导抑制 Bax 诱导的细胞死亡的区域,分别生成Uv1809 N 端(Uv1809Δ246-391)或 C 端(Uv1809Δ23- 245)载体,分别与Bax共浸润烟草发现:编码 Uv1809 C 端部分(Uv1809Δ23-245)载体显示出与全长 Uv1809 相当的抑制 Bax 诱导的细胞死亡的能力(图 1e)。这些结果表明,Uv1809 可能通过干扰植物免疫反应来抑制 Bax 诱导的细胞死亡。
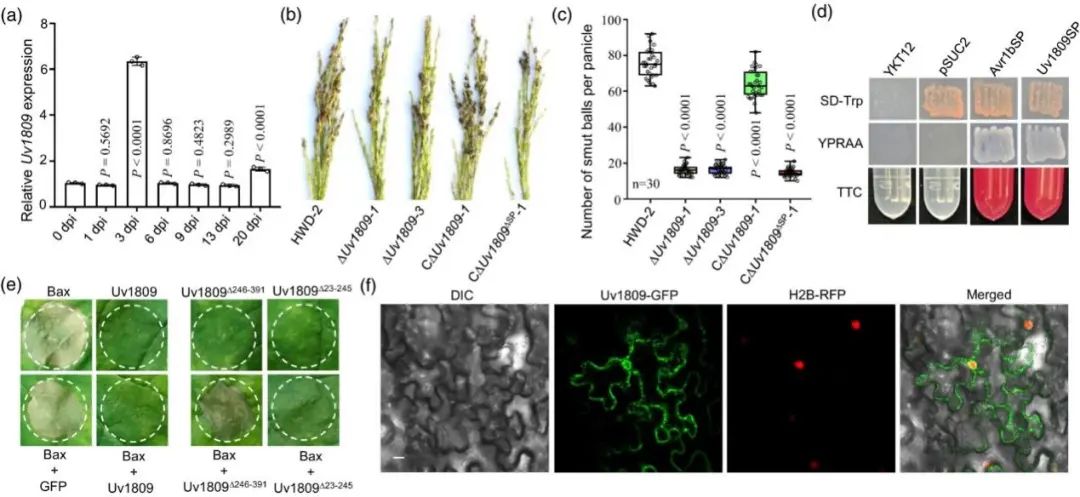
图1. Uv1809是一个关键致病效应因子。
4. Uv1809 是一种细胞内效应器
进一步鉴定Uv1809在烟草叶片中的亚细胞定位。作者构建pCNG-Uv1809 载体导入大肠杆菌EHA105中。与对照不同,表达 Uv1809-GFP 融合蛋白的农杆菌侵染烟草叶片的细胞核和细胞质中检测到 GFP 荧光,但荧光信号会在某些位置聚集(图 1f)。质壁分离后,GFP荧光信号存在于细胞内。Uv1809 是一种细胞内效应物。进一步在RP27 启动子驱动下,将携带核定位信号(NLS)的 Uv1809 与 GFP 融合后异位表达于 Magnaporthe oryzae 菌株 P131 中,脱落大麦叶片接种荧光观测表明Uv1809 可以分泌到植物细胞中。
NLStradamus发现Uv1809 的 aa 336-367 处具有一个潜在的核定位序列 NLS(KDSKDNKDSKANKDSKDNKDSRAVNKPEGKAAR)。亚细胞定位结果表明该 NLS有功能,并成功地将 GFP 定位到了细胞核中。此外,Uv1809Δ246-391 和 Uv1809Δ23-245 在 烟草叶片中亚细胞定位情况表明Uv1809Δ23-245-GFP在细胞核中显示出强烈的 GFP 荧光。因此,Uv1809 的 aa 246-391 在调节 Uv1809 的核定位中起着重要作用。
5. 异源表达 Uv1809 可提高对水稻病原体的敏感性
作者培育异源表达 Uv1809 的转基因水稻株系35S-Uv1809。接种 U. virens 菌株 HWD-2 后,与对照组相比,两个独立的 35S-Uv1809 转基因水稻株系具有更多的稻曲球(图 2a)。35S-Uv1809 转基因水稻株系更易受 Xoo 菌株 PXO99 和水稻稻瘟病菌 M. oryzae 菌株 ZB-25的感染(图 2b,c)。
RT-qPCR表明:防御相关基因 OsPR1b、OsPAL1、OsWRKY13 和 OsAOS2 在 35S-Uv1809 转基因水稻植株中表达显著下调(图2d),这表明 Uv1809 导致了免疫反应的抑制。综上所述,异源表达 Uv1809 可抑制宿主的免疫反应,从而提高水稻植株对病原体的易感性。
6. 通过 Uv1809 的 HIGS 培育抗 RFS 水稻
使用寄主诱导基因沉默技术(HIGS)培育Uv1809 RNAi (Uv1809HIGS)的转基因粳稻。U. virens 侵染表明:与对照水稻植株相比, Uv1809HIGS株系对 RFS 表现出很强抗性(图 2e)。RT-qPCR 定量发现Uv1809HIGS株系中 Uv1809 的相对转录水平远低于对照株系(图 2f)。这一结果表明 Uv1809HIGS 转基因株系对 RFS 的抗性是由 Uv1809 的植物沉默引起的。
为了确定Uv1809是否与Uv1809HIGS株系产生的siRNAs相关,作者对Uv1809HIGS-L1水稻株系进行small RNA测序。测序数据表明:Uv1809HIGS-L1中,与 Uv1809 相匹配的 siRNA丰富,与其相匹配的 siRNA占检测到的所有siRNA 的 0.83%。想匹配的 siRNA 大小在 18-30 个核苷酸之间, 21-核苷酸的 siRNA 数量最多(图 2g)。荧光原位杂交显示水稻花组织和 U. virens 感染的菌丝中具有荧光信号(图 2h)。这表明 Uv1809-RNAi 载体能在 Uv1809HIGS-L1水稻株系中产生有效的针对 Uv1809 的 siRNA,在感染过程中转运到真菌细胞中,从而降低入侵的 U. virens 菌丝中的 Uv1809 转录水平。
综上所述,在转基因水稻植株中沉默 Uv1809 能增强对 U. virens 的抗性, Uv1809 是感染过程中的一个关键致病性效应因子。
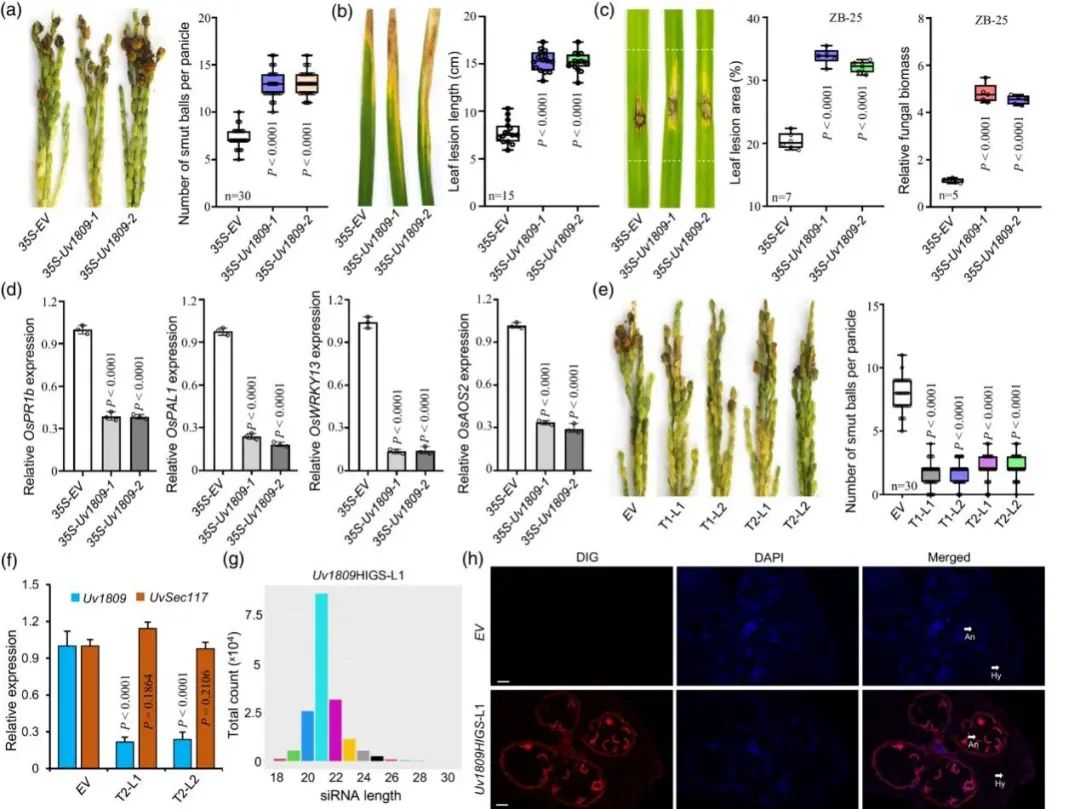
图2. Uv1809异源过表达提高水稻对病原菌的敏感性,Uv1809的HIGS增强了水稻对稻瘟病菌的抗性
7. Uv1809 与水稻组蛋白去乙酰化酶 OsSRT2 相互作用
提取U. virens 感染的水稻小穗RNA构建 cDNA 文库。Uv1809作为biat进行酵母双杂交筛选。结果表明水稻组蛋白去乙酰化酶 OsSRT2 是 Uv1809 的靶标,且 Uv1809 与全长 OsSRT2 互作(图 3a)。瞬时共浸有 OsSRT2-Flag 和 Uv1809-GFP 的烟草叶片的Co-IP(体内互作)发现Uv1809 被 OsSRT2 免疫沉淀(图3b)。GST pull down 实验表明Uv1809ΔSP-His 和 OsSRT2-GST 在体外互作(图3c)。Uv1809-cYFP 和 OsSRT2-nYFP 融合载体瞬时共浸润烟草叶片,双分子荧光互补(BiFC)分析发现烟草叶表皮细胞细胞核中观察到了 YFP 荧光(图 3d)。此外,Uv1809 片段(Uv180923-245 和 Uv1809246-391)中Uv1809246-391与 OsSRT2互作。
因此,Uv1809在植物细胞核中与OsSRT2相互作用,uv 1809246–391是与OsSRT2相互作用的关键区域。
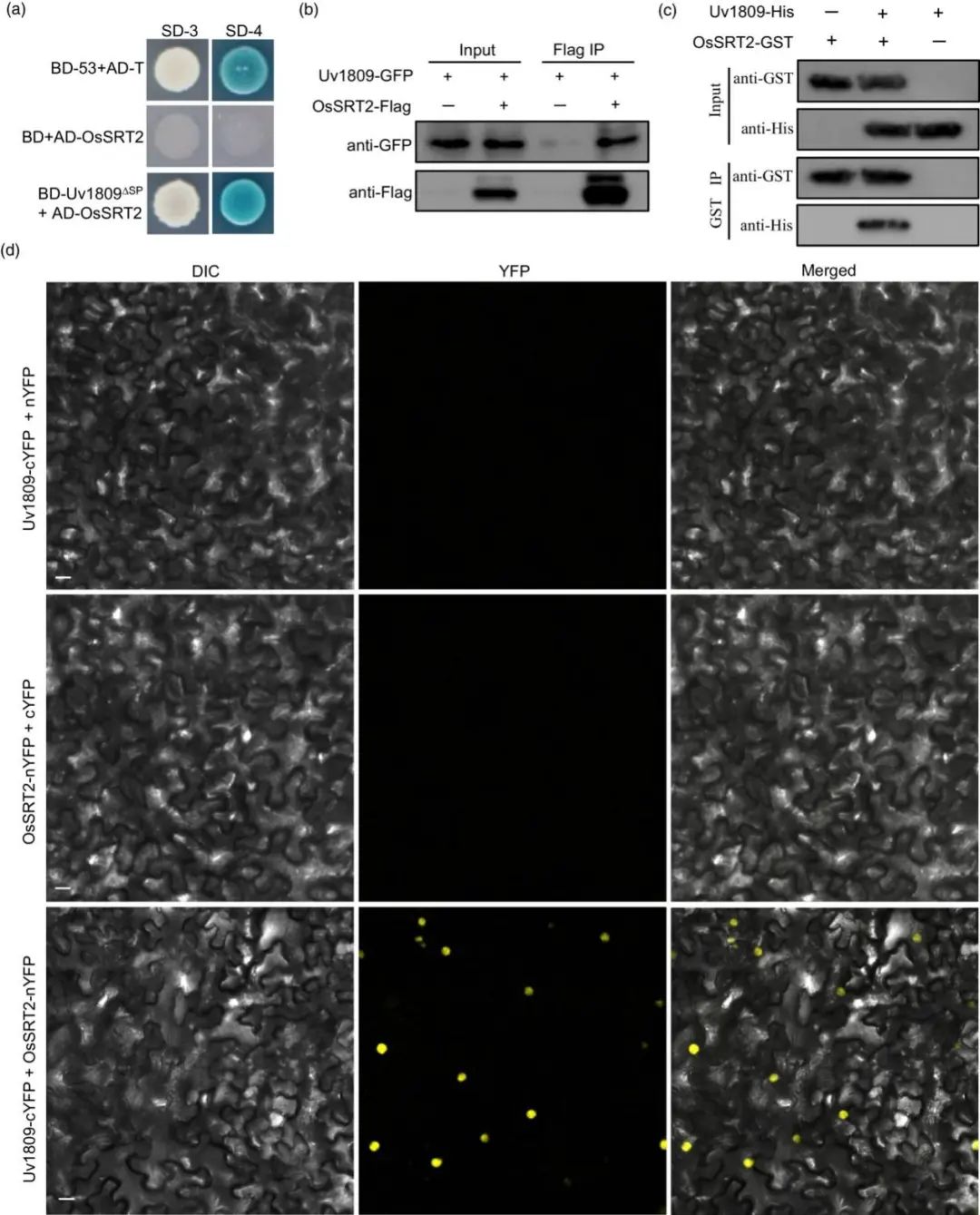
图3. Uv1809与OsSRT相互作用。
8. OsSRT2 负调控水稻对病原体的广谱抗性
首先RT-qPCR 数据表明U. virens感染过程中,OsSRT2 的转录水平显著上调。因此,OsSRT2 的表达受到 U. virens 感染的诱导,OsSRT2 可能对水稻对 U. virens 的抗性有负调节作用。
烟草亚细胞表明 OsSRT2 定位于细胞核,使用CRISPR/Cas9技术编辑产生OsSRT2 基因敲除突变株系(ossrt2)。ossrt2水稻株系形态与野生型ZH11相似(图4a)。分别接种 U. virens 菌株 HWD-2 的菌丝体/孢子悬浮液、稻瘟病菌 M. oryzae 菌株 P131 或 ZB-25、Xoo 菌株 PX099和R. solani 菌株 HG81后ossrt2:的稻曲球远少于 ZH11 株系(图 4b);对 M. oryzae 具有很强的抗性(图 4c-f);P131产生的病变更短(图 4g);对 HG81具有更好的抵抗力(图 4h)。此外,接种 U. virens 1 dpi,RT-qPCR显示ossrt2中防御相关基因(OsPR1b、OsPR10a、OsPAL1 和 OsSGT1)转录水平显著增加(图 4i)。
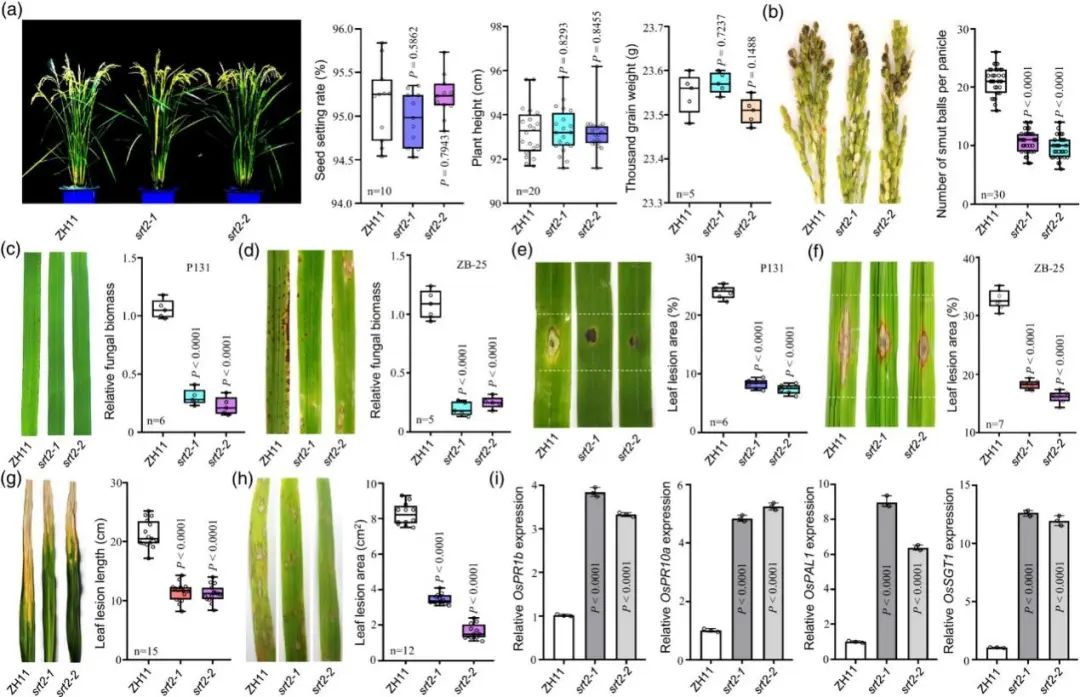
图4. OsSRT2负调控水稻对病原菌的广谱抗性。
9. Uv1809 提高 OsSRT2 的去乙酰化活性
水稻 HDAC 可调节组蛋白 H3 或 H4 的乙酰化水平。作者进一步研究OsSRT2 是否影响组蛋白去乙酰化。从ossrt2 和 ZH11 水稻小穗中分离组蛋白,广谱的抗H3ac 和抗 H4ac 抗体免疫印迹分析发现ossrt2 突变体中 H4K5ac 和 H4K8ac水平升高,H3上水平无增加(图 5a)。因此,OsSRT2 调节水稻小穗中 H4K5 和 H4K8 的组蛋白乙酰化水平。
此外,35S-EV 和 35S-Uv1809 水稻稻穗中 H3ac 和 H4ac 的水平检测发现35S-Uv1809水稻株系中的H3K9ac、H4K5ac和H4K8ac水平明显降低(图5b),这表明Uv1809可能通过影响OsSRT2来抑制水稻的H4K5ac和H4K8ac。
进一步检测重组OsSRT2-GST单独或在 Uv1809ΔSP-His、Uv180923-245-His 或 Uv1809246-391-His 存在下的活性。OsSRT2-GST 与 Uv1809ΔSP-His 或Uv1809246-391-His 一起孵育,去乙酰化活性增加(图5c)。OsSRT2介导的 H4K5ac 和 H4K8ac 的脱乙酰化通过与重组 Uv1809ΔSP-His 或 Uv1809246-391-His 共孵育而增强(图5d)。烟草叶片中瞬时表达 OsSRT2-Flag、 OsSRT2-Flag 和 Uv1809-GFP、 OsSRT2-Flag 和 GFP 以及 Uv1809-GFP,核蛋白提取以及H4K5 和 H4K8 乙酰化水平评估发现,OsSRT2-Flag 和 Uv1809-GFP 共同表达降低了组蛋白 H4K5 和 H4K8 的乙酰化水平(图 5e)。因此,OsSRT2-Flag 和 Uv1809-GFP 共表达增加核蛋白的去乙酰化活性(图 5f)。
综上所述, Uv1809 在体外和体内都能增强 OsSRT2 介导的组蛋白去乙酰化作用。
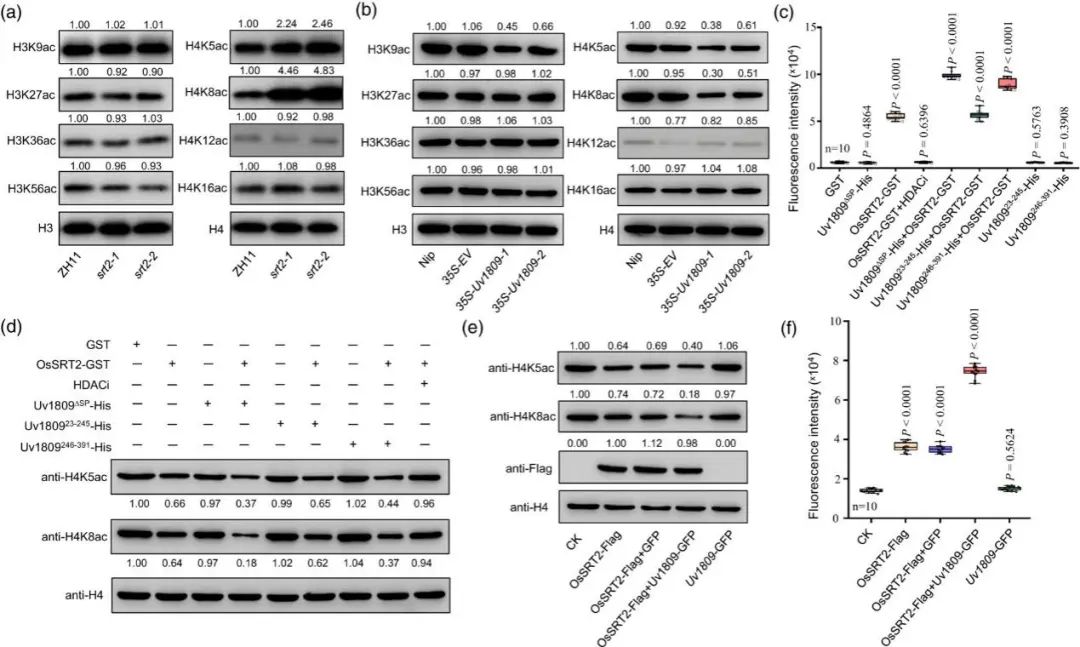
图5. Uv1809对OsSRT2组蛋白去乙酰化活性的影响。
10. OsSRT2 通过调节 H4K5ac 和 H4K8ac 标记基因抑制水稻免疫力
对野生型 ZH11和ossrt2水稻小穗进行RNA测序,数据分析表明ossrt2 中分别有 604(上调) 和 378(下调)个差异基因(图 6a)。基因集富集分析(GSEA)表明,真菌防御反应调控和防御反应通路的调控被显著富集,这些通路中的基因在ossrt2中被上调(图6b)。ossrt2中显著上调基因GO富集分析显示防御反应调控、茉莉酸介导信号通路调控和刺激反应调控通路被富集(图 6c)。因此,OsSRT2 突变可能会促进水稻小穗防御途径,并赋予这些突变体对病原体感染的抗性。
ChIP-seq进一步研究OsSRT2 在感染过程中的基因调控机制。比较野生型和ossrt2 植株小穗中 H4K5ac 和 H4K8ac 的全基因组图谱。ChIP-seq数据分析表明ossrt2中共发现了 657 个 H4K5ac 水平升高的峰(458 个基因)和 316 个 H4K5ac 水平降低的峰(249 个基因)(图 6d)。H4K8ac 比较分析表明,ossrt2 中共有 1236 个峰(932 个基因)的 H4K8ac 沉积显著增加,而 srt2 中只有 74 个峰(61 个基因)的 H4K8ac 沉积大幅减少(>1.5 倍,P < 0.05)(图 6d)。这些发现与之前ossrt2 中 H4K5ac 和 H4K8ac数据一致(图 5a)。
H4K5ac和H4K8ac峰(基因)重叠分析结果显示,ossrt2中58.4%(61.2%)的H4K5ac上调峰(基因)与H4K8ac上调峰(基因)重叠(图6e),暗示ossrt2对H4K5ac和H4K8ac沉积具有类似影响。GO富集分析显示62 个 GO terms与 H4K5 和 H4K8 高乙酰化基因相关,涉及包括防御相关途径在内的多种生物过程(图6f)。
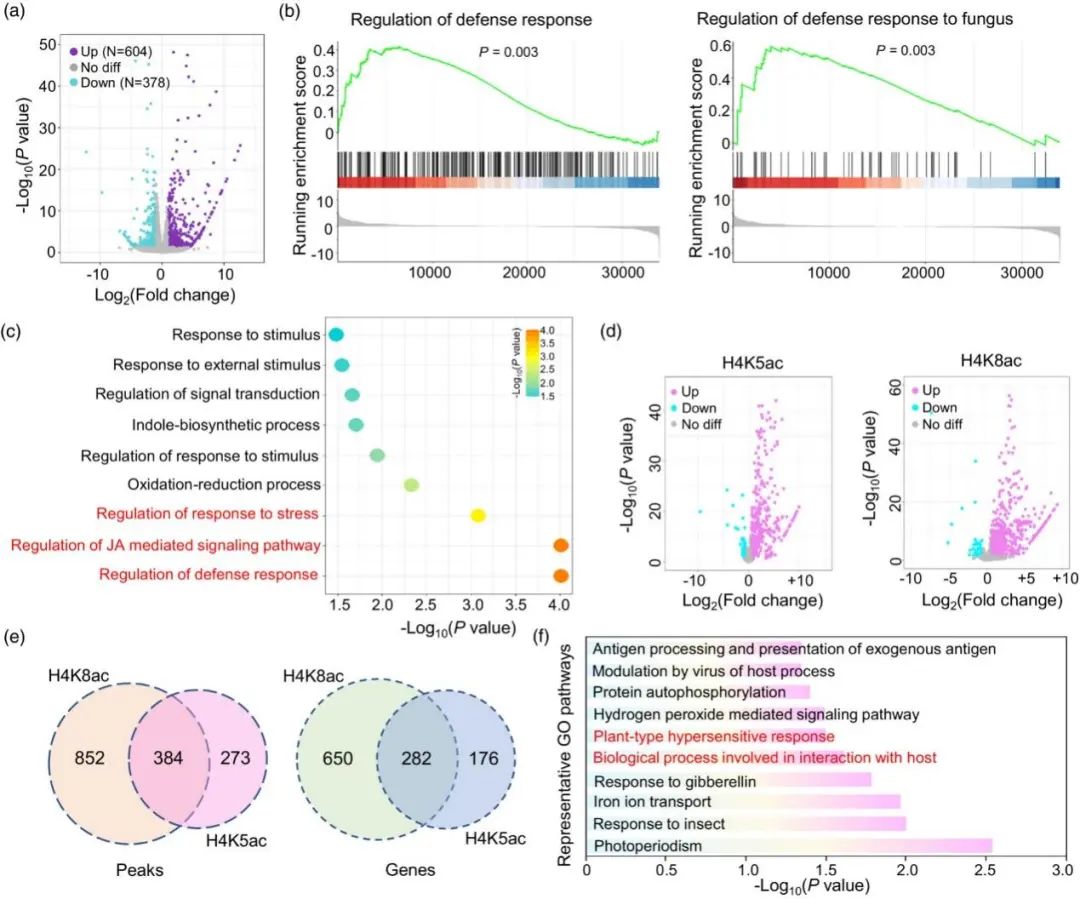
图6. Ossrt2和野生型水稻小穗的RNA-seq和CHIP-seq数据分析。
为了研究水稻小穗中转录组和组蛋白乙酰化之间的关系,作者对H4K5/8ac的动态变化和ossrt2中基因转录之间的相关性进行检测。发现在ossrt2中H4K5/8ac和基因表达变化之间具有中等水平的正相关 (图7a)。ossrt2中,84个基因呈现转录上调和H4K5/8ac修饰超乙酰化(图7c)。GO富集分析鉴定出56个GO terms,其中植物防御相关途径是显著的,包括高渗反应、先天免疫反应激活和植物型超敏反应(图7d)。ChIP-qPCR分析表明ossrt2水稻防御相关基因启动子区域组蛋白上H4K5ac或H4K8ac水平的增加导致转录更活跃(图7e)。
总之, OsSRT2对H4K5/8的去乙酰化抑制了植物防御相关基因的转录,导致水稻免疫下降。
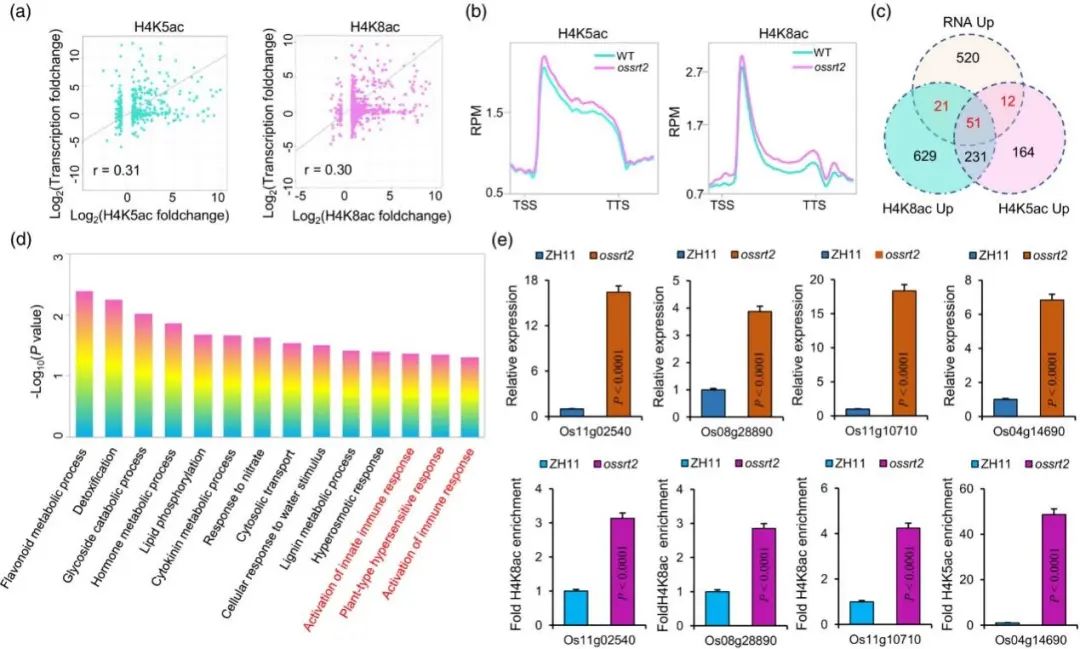
图7. H4K5ac和H4K8ac在ossrt2水稻小穗基因表达调控中的作用。
# 总 结 #
该研究证明分泌型蛋白Uv1809是一个关键致病性因子。Uv1809靶向增强水稻组蛋白脱乙酰酶OsSRT2介导组蛋白脱乙酰化,降低H4K5ac和H4K8ac水平,从而干扰防御基因的转录激活,进而抑制水稻免疫。此外由CRISPR-Cas9技术编辑的ossrt2水稻株系在生长和产量方面无不良影响,但对水稻病原菌表现出广谱抗性,这为抗病育种提供了潜在有价值的遗传资源。
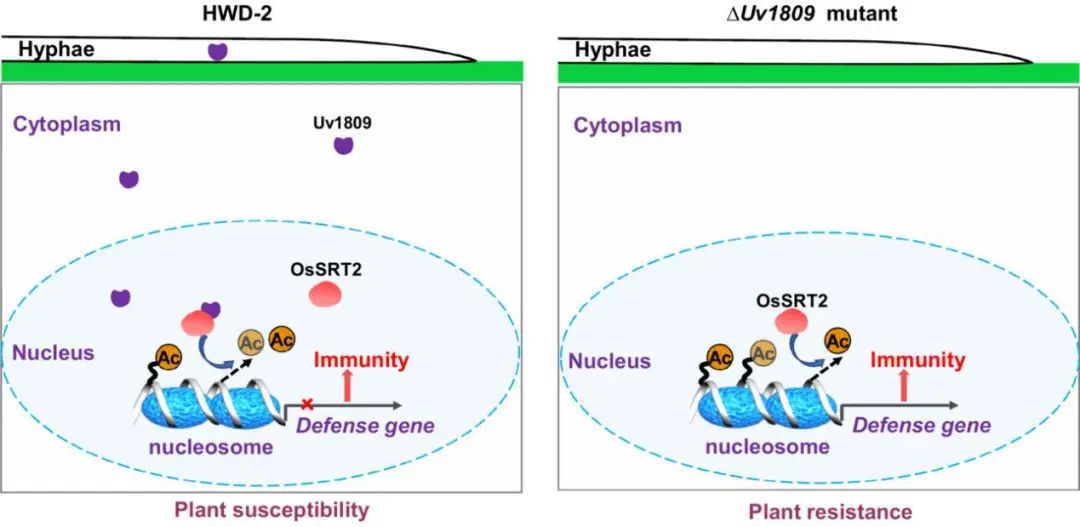
图8. 模式图
原文链接:https://doi.org/10.1016/j.celrep.2023.112688
# 关于我们 #
爱基百客专注于提供领先的表观组学服务,可提供方案设计、样本制备、测序、分析以及验证一站式服务。
九月开学季,爱基百客带着表观产品前来助力!活动期间,表观产品全线优惠,部分产品低至8折!活动即将结束,有表观产品需求的老师,不要错过啦~
项目咨询WX:Igenebook0
这篇关于项目文章| PBJ(IF:13.8)发表稻曲病菌效应因子Uv1809增强组蛋白去乙酰化抑制水稻免疫的分子机制的文章就介绍到这儿,希望我们推荐的文章对编程师们有所帮助!




